Weichtiere
aus dem Rheinischen Mitteldevon
Von Hans Josef Jungheim
Attraktive Funde werden immer
seltener
Neben den Stachelhäutern, zu
denen die Seelilien zählen, und den Gliederfüßern mit
den Trilobiten liefern die Weichtiere besonders begehrte Fossilien
aus dem Mitteldevon der Eifel, zumal auch hier attraktive Funde
seltener werden, obwohl die Vielzahl der aufgesammelten Fragmente
durchaus für den Arten- und Individuenreichtum einzelner
Weichtiergattungen spricht.
Der formenreiche Stamm der
Weichtiere gliedert sich in fünf Klassen, von denen die Muscheln
(Lamellibranchiata), die Kopffüßer (Cephalopoda) und die
Schnecken (Gastropoda) die bekanntesten sind. Fossile Vertreter sind
überall im Rheinischen Mitteldevon aufzufinden.
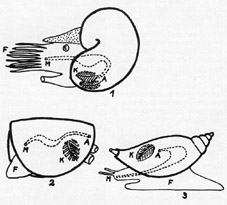
Weichkörper
und Gehäuse (dicke Linien) von Mollusken: 1 = Kopffüßer,
2 = Muschel, 3 = Schnecke; F = Fuß, A = After, K = Kiemen, M =
Mund (nach R. C. Moore, 1952). Der ungegliederte Körper der
Mollusken zeigt i. d. R. vier Regionen: Kopf mit Augen und Fühlern
- muskulöser Fuß - Eingeweidesack - Mantel, d. i. eine in
der Rückenregion liegende Hautfalte, die u. a. die Schale
absondert.
Die Weichtiere, zu denen noch die
Käferschnecken und Grabfüßer zu zählen sind,
unterscheiden sich von Gruppe zu Gruppe sowohl nach Körperbau
als auch nach Lebensweise. Sie besitzen keinen einheitlichen Bauplan.
Sie haben im Verlauf ihrer Entwicklung einen sogenannten Mantel
ausgebildet, der einmal Teile des Körpers bedeckt und zum andern
die Hartteile, wie Schale oder Gehäuse aus Kalziumkarbonat
(CaCO3) absondert. Letzteres gilt allerdings nicht mehr
für alle Mollusken, von denen ein Teil auf das ursprünglich
bei allen Weichtieren vorhandene Grundmerkmal, nämlich der
Besitz einer die Weichteile nach außen hin schützenden
Schale, inzwischen verzichtet.
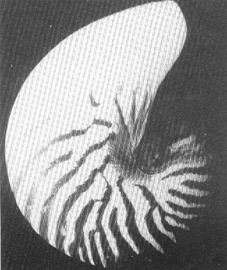
NAUTILUS
POMPILIUS L., Ansicht auf das Gehäuse von außen. Die
Schale ist glatt und weist eine einfach gewellte Lobenlinie auf. Der
Nabel ist ziemlich eng. N. POMPILIUS frißt Korallen und hält
sich darum vornehmlich am Boden der Korallengründe auf.
(Durchmesser auf Nabelhöhe = 12.2 cm.)
Ohne Zweifel zählen die
Mollusken, die zahlreiche Fossilien liefern, zu den wichtigsten und
erfolgreichsten Tierstämmen. Rezent sind nahezu 130.000 Arten,
fossil seit dem Kambrium über 10.000 Gattungen bekannt. Der Name
leitet sich von (lat.) mollis = weich ab; er bezieht sich auf den
weichen, sklettlosen aber vielgestaltigen Körperbau der
„Weich“-tiere, für die ein ventraler „Fuß“
und der von der Dorsalseite ausgehende „Mantel“ typisch
ist.
Die meisten Weichtiere besitzen
einen gestreckten, im ganzen auch zweiseitig symmetrischen Körper
mit deutlich abgesetztem Kopfabschnitt (außer bei den Muscheln)
und gut entwickelten Sinnesorganen. Sie verfügen u. a. über
ein Blutgefäßsystem, einen Herzbeutel und
Geschlechtsdrüsen. Neben winzigen Formen existieren riesige
rezente Weichtiere, unter denen die in der Tiefsee lebenden
Riesenkalmare mit einer Länge bis nahezu zwanzig Metern
(einschließlich der Greifarme) die größten
wirbellosen Tiere darstellen.
Weichtiere sind in der Regel - von
den Kopffüßern einmal abgesehen - nur wenig
bewegungsfreudig. Die meisten leben festsitzend oder kriechend. Wenn
die Mehrzahl aller Weichtierarten auch nach wie vor im Meer lebt, so
hat doch zum Beispiel eine Reihe von Schneckenarten neben
Süßwasserbereichen auch das Festland erobert. Während
die Muscheln bis in brackiges und süßes Wasser vordringen
konnten, bleiben die Kopffüßer ganz auf das Leben im Meer
beschränkt.
Für das Rheinische
Mitteldevon sind Muscheln, Kopffüßer und Schnecken von
besonderer Bedeutung. Vor allem der Artenreichtum der Schnecken ist
durchaus bemerkenswert. Bei einigen Muschelarten fällt das
massenhafte Vorkommen der Individuen auf, die oft ganze Gesteinsbänke
durchsetzen. Funde von Cephalopodengehäusen sind inzwischen
seltener zu erwarten; Bruchstücke lassen sich allerdings an
einigen wenigen Fundorten immer wieder finden.
Muscheln
Die Muscheln (Lamellibranchiata)
besitzen eine zweiklappige Schale (Bivalvia). Sie werden darum häufig
vom Laien mit den ebenfalls zweiklappigen Brachiopoden verwechselt.
Der größte Teil aller
Muscheln lebt grabend in den obersten Partien des Meeresbodens mehr
oder weniger standorttreu. Manche Arten graben sich bis zu 1,50 m
tief ein. Hierzu bedurfte es allerdings in der Evolution u. a. des
Erwerbs besonders leistungsfähiger Kiemenpumpen und Siphone, um
sich einen neuen, bis dahin ungenutzten Biotop, hier das Sediment -
Innere, zu erschließen. Andere Muscheln können durch das
rhythmische Öffnen und Schließen ihrer beiden Klappen frei
im Wasser schwimmen. Sie zeigen hierbei allerdings wenig Ausdauer und
liegen zumeist seitlich auf dem Meeresboden, während bei den
grabenden Muschelarten die Schalen senkrecht stehen. Ein weiterer
Teil von Muschelarten ist am jeweiligen Untergrund festgewachsen.
Wieder andere besitzen die Fähigkeit, sich in Gestein, und in
Holz einzubohren. Diese bohrenden Muscheln benutzen hierzu entweder
eine scharfätzende Säure, die aus besonderen Drüsen
abgesondert wird, oder aber einen sogenannten Raspelbohrer (scharfe
Zähnchen besetzen die vorderen Kanten der Schale, mit der die
Bohrmuschel eine Drehbewegung ausführt und auf diese Weise das
Gestein abschleift bzw. zerraspelt).
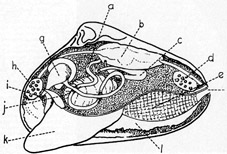
Schnitt
durch eine Muschel
a = Darm, b = herz, c = Niere, d = hinterer
Schließmuskel, e = Kiemen, f = Siphone (Aus- und
Einströmsipho), g = Magen, h = vorderer Schließmuskel, i =
Mundöffnung, j = Eierstock, k = Fuß, l = Mantel.
Die meisten Muscheln zählen
zu den Benthonten, d. h. sie leben am Grund von Gewässern. Darum
sind für sie schützende Außenskelette besonders
wichtig.
Die Muscheln sind kopflose
Weichtiere, die ausschließlich im Wasser leben. Zwei fleischige
Mantellappen scheiden die beiden kalkigen Klappen der zumeist ovalen
oder, wie etwa bei den Bohr- und Miesmuscheln, quer verlängerten
Schale aus, die hauptsächlich dem Schutz der Weichteile dient
aber auch andere Funktionen übernimmt. Die Weichteile bestehen
im wesentlichen neben den Mantellappen noch aus einem Paar Kiemen,
dem Rumpf mit den Eingeweiden und einem muskulösen Fuß,
der in der Regel zum Kriechen oder Graben dient. Er kann stets ganz
in das Innere der Schale zurückgezogen werden. Wenn die Muscheln
auch keinen Kopf besitzen, so fehlen ihnen doch nicht die für
die anderen Weichtierklassen typischen Sinnesorgane. Viele Arten
verfügen über zahlreiche Augen, Tastfäden und
Geschmacksorgane. Die Kiemen, über die die Atmung erfolgt,
liegen unter den Mantellappen.
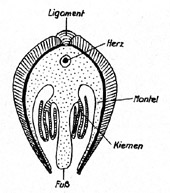
Schematischer
Querschnitt durch eine Muschel (nach A. H. Müller, 1965). Zwei
fleischige Mantellappen umhüllen das Muscheltier, dessen Atmung
über die Kiemen erfolgt, die sich unter dem Mantellappen
befinden. Das Herz umschließt scheidenartig den Enddarm. Die
Muschelschale besteht i. d. R. aus zwei verschiedenen Schichten: a)
die aus Calcit bestehende Prismenschicht (Ostrakum) außen, b)
die Perlmutterschicht (Hypostrakum) innen.
Die Muschelschale besteht aus zwei
Klappen, die durch das elastische Ligament unterhalb des Wirbels
zusammengehalten und durch Muskeln (Schließmuskeln) geschlossen
werden. Die Mehrzahl aller Muscheln verfügt über ein Schloß
mit Schloßzähnen und den entsprechenden Vertiefungen
(Zahngruben). Das Schloß sichert den Halt der beiden
Schalenhälften. Die Ausbildung des Schloßverbandes ist
unterschiedlich, so zum Beispiel nach der Anzahl und Anordnung der
Zähne und Zahngruben und ihrer Größe. Die acht
Hauptschloßtypen besitzen eine große Bedeutung für
die Systematik der Lamellibranchiata.
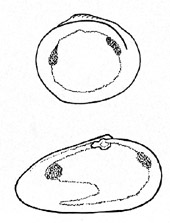
Schematische
Darstellung über die Ausbildung der Schließmuskeleindrücke
und den Mantellinienverlauf bei Muscheln (nach A. H. Müller,
1965). Zwei von sechs verschiedenen Möglichkeiten: Oben eine
Form ohne, unten mit Mantelbucht. Die Mantellinie oben zeigt einen
einfachen, nicht unterbrochenen Verlauf.
Die Muscheln haben sich im Verlauf
ihrer stammesgeschichtlichen Entwicklung mannigfaltig umgestaltet, so
daß es schwierig ist, allein mit Hilfe der heute lebenden Arten
eine natürliche Systematik aufzustellen. Erschwerend kommt
hinzu, daß etwa ein Viertel der und siebzig Familien erst mit
der Kreidezeit auftritt und ein Dutzend Familien keine rezenten
Vertreter aufweist. Leider sind die aus dem älteren Paläozoikum
stammenden Vertreter in der Regel nur schlecht erhalten, weil ihre
Schalen vornehmlich aus Aragonit (CaCO3) bestanden, der
leicht löslich ist. Hieraus ergeben sich für taxionomisch
Klärungen zusätzliche Schwierigkeiten; denn ein
geschlossenes System fordert die Verknüpfung der fossilen mit
den rezenten Organismen.
Das Wachstum der Muschelschale
beginnt - wie bei der Brachiopodenschale - jeweils am Wirbel jeder
der beiden Klappen. Die Klappen wachsen in der Regel durch eine
einfache Anlagerung von Skelettmaterial. Die Anwachslinien auf der
Schalenaußenfläche kennzeichnen die Wachstums-
(ontogenetischen) Stadien. Die Muschelschale gibt Auskunft nicht nur
über die zahl und die Größer er Schließmuskeln
(die Muskelanwachsstellen bleiben auch bei fossilen Exemplaren oft
sichtbar), sondern kennzeichnet über den Verlauf der Mantellinie
Form und Ausdehnung der Mantellappen. Hierdurch sind wichtige
Bestimmungsmerkmale auch bei fossilen Formen fixiert.

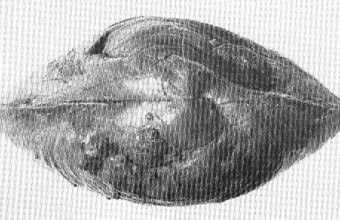
Paracyclas
proavia
(Ansicht von der Seite)
Die zumeist dicke Schale
zeigt i. d. R. und bei guter Erhaltung konzentrische Anwachsstreifen.
Die Muschel ist durchweg breiter als lang. (Breite des vorliegenden
Exemplars 6,2 cm.)
FO: Sötenich, Eifel, Wachtberg,
Mittel-Devon.
(Ansicht
auf das Schloß)
Muscheln, von denen fast 300
Gattungen fossil belegt sind, existieren bereits im späten
Kambrium, werden allerdings erst mit dem Ordovicium etwas häufiger,
treten aber bis zum Ende des Erdaltertums (Perm) insgesamt wenig
hervor, zählen jedoch neben den Brachiopoden zu den häufigsten
permischen Fossilien. Lediglich im Devon werden sie erstmalig
zahlreicher und füllen im Mitteldevon, wenn auch mit wenigen
Arten, Kalksteinbänke und Riffpartien auf (vor allem Paracyclas
und Megalodeon). Im Oberkarbon nehmen Süßwassermuscheln
kurzfristig zu.

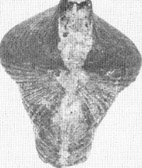
Megalodon
abbreviatus
(Ansicht von vorn auf die Schloßregion)
Bei
M. abbreviatus handelt es sich um eine relativ große,
gleichklappige Muschel mit dicker, oft glatter oder auch konzentrisch
gestreifter, gewölbter Schale mit ovaler Form. Die kräftigen
Wirbel der beiden Klappen sind gebläht und nach vorne
eingekrümmt. Bei ausgewachsenen Exemplaren ist die Länge
größer als die Breite. (Länge des abgebildeten
Exemplars 7,7 cm.)
FO: Bergische Gladbach, Schlade, Bücheler
Schichten, Ob. Mittel-Devon, Bergisches Land
Foto: Kreisbildstelle
*)
Mit dem Beginn des Erdmittelalters
gewinnen die Muscheln zunehmend an Bedeutung, liefern ab der Trias
auch zahlreiche wichtige Leitfossilien und erleben in der Kreidezeit
(Oberkreide) ihr absolutes Maximum. Besonders bemerkenswert ist
hierbei das erstmalige Auftreten riffbildender Lamellibranchiata,
wobei vor allem die Rudisten als Ergebnis fortschreitender Anpassung
an Riffbedingungen korallenähnliche Wuchsformen entwickeln.
Conocardium
clathratum
Eine kleine, gleichklappige Muschel mit verlängert
dreieckigem umriß, deutlich radial berippt. Die Schloßseite
ist abgestutzt und seitlich verlängert. Sie zeigt einen langen
und geraden Schloßrand. (Länge 1,4 cm.)
FO: Bergisch
Gladbach, Schlade, Bücheler Schichten, Ob. Mittel-Devon,
Bergisches Land
Foto: Kreisbildstelle *)
Nach der Kreidezeit erfahren die
Muscheln einen allerdings nur schwachen Rückgang bis zur
Jetztzeit, während beispielsweise die ebenfalls zu den Mollusken
zählenden Schnecken vom Tertiär an deutlich hervortreten
und die Muscheln an Bedeutung übertreffen.
Im Rheinischen Mittel-Devon
ralativ häufig vorkommende Muschelarten sind zum Beispiel
Megalodon abbreviatus, M. Cucullatus, Paracyclas proavia und
Conocardium clathratum.
Kopffüßer
Die Kopffüßer leben
ausschließlich im Meer. Bis auf den Nautilus, der im
südwestlichen Teil des Pazifischen Ozeans rezent anzutreffen
ist, sind alle übrigen gehäusetragenden Vertreter dieser
hochentwickelten Tiergruppe ausgestorben.
Die sonst noch heute lebenden
Formen, wie Kalmare und Kraken, besitzen kein eigentliches Gehäuse,
sondern nur mehr eine hornartige Platte tief in der Haut.
|
Tetrabranchiata |
Nautiloidea
Ammonoidea |
vierkiemige
Kopffüßer |
|
Dibranchiata
|
Belemnoidea
Sepiodea
Teuthoidea
Octopoidea |
zweikiemige
Kopffüßer |
Die besonderen Kennzeichen der
Cephalopoden sind einmal der Trichter, der als Öffnung der
Mantelhöhle u. a. als Schwimmorgan dient, und die am Kopf
sitzenden Tentakeln (Greif- und Bewegungsorgane), die in der
Hauptsache für den Erwerb der Nahrung sorgen. Der Kopf verfügt
zudem über zwei Augen und, ganz im Gegensatz zu den übrigen
Mollusken über ein insgesamt gut entwickeltes Gehirn.
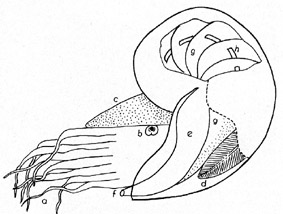
Medianschnitt
durch den rezenten Cephalopoden NAUTILUS POMPILIUS L.
(vereinfachte Darstellung): a = Tentakeln, b = Auge, c = Kopfkappe, d
= Kieme, e = Mantellappen, f = Trichter, g = Wohnkammer, g =
Luftkammern, h = Sipho.
Die Kopffüßer sind
nicht nur die höchstentwickelten Weichtiere, sie stellen auch
die größten bekannten Tierformen unter den Wirbellosen.
Die rezente Gattung Architeuthis erreicht zum Beispiel eine
Länge von 6,5 Metern.

„Belemniten-Haufwerk“
mit DACTYLOTEUTHIS, SALPINGOTHEUTHIS u. a., Jura, Lias Zeta
(Ob. Toarcium).
FO: Mistelgau b. Bayreuth.
Die Belemniten sind
ausgestorbene dibranchiate Cephalopoden, von denen i. d. R. nur die
massiven Teile (zumeist, wie hier, die Rostren = „Donnerkeile“)
fossil gefunden werden.
(Länge der Platte 18 cm.)

Längsschnitt
durch das Gehäuse des rezenten Cephalopoden NAUTILUS
POMPILIUS L. Deutlich sichtbar sind die relativ einfach gebogenen
Kammerscheidewände voneinander getrennten Kammern, die von der
sogenannten Siphonalröhre durchzogen werden.
Alle Kopffüßer sind
ausgezeichnete Schwimmer, die sich mit Hilfe eines
Rückstoßmechanismus - sie pressen einen kräftigen
Wasserstrom aus ihrem muskulösen Mantel - sehr schnell nach
allen Richtungen fortbewegen können.

Ammoniten-Gehäuse
aus der Jurazeit (Erdmittelalter).
DACTYLIOCERAS COMMUNE
(FO: Schlaifhausen).
Das Cephalopodengehäuse
(Phragmakon) ist im Gegensatz zum Schneckengehäuse gekammert.
Das eigentliche Tier lebt nur in der äußersten und zuletzt
gebildeten Kammer, der sog. Wohnkammer, die den nach vorne offenen
Teil des Cephalopodengehäuses darstellt und in den sich das Tier
bei Gefahr zurückziehen kann. Die übrigen Kammern sind mit
Gas gefüllt. Durch die Zwischenwände (Kammerscheidewände),
auch Septen genannt, führt ein Kanal, der Sipho, der alle
Kammern miteinander verbindet und mithilft, den Gasdruck zu
regulieren. Die Gasfüllung verringert das spezifische Gewicht
des Gehäuses, ohne dessen Schutzfunktion zu beeinträchtigen.
Der Sipho ist ein besonders charakteristisches Organ der Kopffüßer.
Es handelt sich hierbei um einen kalkigen, mehr oder weniger
zylindrisch geformten Strang, der von der Wohnkammer aus durch alle
Kammern des Gehäuses reicht und je nach Gattung und Art zentral
oder peripher verläuft.

Teilstück
eines Siphos von Cyrtoceras. (Länge 3,3 cm.)
FO: Sötenich,
Eifel, Mittel-Devon.
Foto: Kreisbildstelle *)
Die Gestalt des
Cephalopodengehäuses ist außerordentlich variabel. Neben
gestreckten kommen gebogene oder spiralig aufgerollte Gehäuseformen
vor. Auch die Struktur der Schalenoberfläche zeigt einen
beträchtlichen Variationsreichtum.
Nach der Anzahl ihrer Kiemen
werden die Kopffüßer in zwei Unterklassen eingeteilt:
Für das Devon sind
ausschließlich die vierkiemigen Cephalopoden von Bedeutung. Die
Dibranchiata treten erstmalig mit den Belemniten, die sich
wahrscheinlich aus den Nautiloideen entwickelt haben, im Karbon auf.
Die ältesten fossilen Cephalopoden dürften wohl bis in das
späte Kambrium zurückreichen. Im Ordovicium, also noch im
frühen Erdaltertum, treten sie plötzlich mit großer
Formenfülle auf, wobei möglicherweise sogar die ersten und
noch stabförmigen Ammonoideen erscheinen. Während die
Nautiloideen schon zum Beginn der Devonzeit an Bedeutung verlieren,
erlebt der Zwei der Ammonoideen eine um so reichere Entfaltung; ihre
bis dahin gestreckten Gehäuse beginnen sich nun einzurollen. Die
Goniatiten liefern für das Oberdevon ausgezeichnete
Leitfossilien. Mit dem Beginn des Erdmittelalters (Trias) werden die
Ammoniten zahlreicher und bevölkern mit vielen Gattungen die
Ozeane. Wenn sich von den Nautiloideen vom Tertiär an nur mehr
der bereits mehrfach erwähnte NAUTILUS bis heute erhalten
hat, so sind die Amonoideen gänzlich ausgestorben. Sie treten
zwar bereits im Ordovicium mit den ältesten Formen auf,
überflügeln im späten Erdaltertum die Nautiloideen,
übernehmen schließlich im Mesozoikum mit ungeheurer Arten-
und Individuenzahl bis zum Ende der Kreidezeit die Vorherrschaft
unter den Kopffüßern, um dann aber aus bisher ungeklärter
Ursache in kurzer Zeit vollständig zu verschwinden. Bereits vor
Beginn der Jurazeit sterben die Nautiloideen, Ammonoideen und
Belemnoideen bis auf je eine relativ einfache gebaute Stammform, die
noch einmal neue differenzierte Entwicklungen bis in die Kreidezeit
einleiten, aus. Die gehäusetragenden Cephalopoden treten bis auf
NAUTILUS ab. Die Klasse der Cephalopoden entwickelt nun
Formen, die ihre kalkige Schale als Schulp nach innen verlagern.

Comphoceras
sp.
Ein durch Verwitterung beschädigtes Exemplar aus der
Bergisch Gladbach-Paffrather Kalkmulde. Die Luftkammern liegen z. T.
offen, die Wohnkammer fehlt. (Länge des Fossils 5,3 cm.)
Foto:
Kreisbildstelle *)

Cyrtoceras
depressum
Das im Querschnitt runde Gehäuse ist nur wenig
eingekrümmt (cyrtoceroid) und verjüngt sich in seinem
hinteren Teil sehr rasch. Die einfach gebogenen Kammerscheidewände
folgen dicht aufeinander. Der zumeist perlschnurartig ausgebildete
Sipho liegt an der inneren Peripherie. (Länge des
unvollständigen Exemplars 28,0 cm.)
FO: Sötenich, Eifel,
Mittel-Devon.
Foto: Kreisbildstelle *)
Es ist mehr als wahrscheinlich,
daß die ältesten Ammoniten während eines relativ
kurzen Zeitabschnittes des Unterdevons im Bereich des heutigen
Hunsrücks entstanden sind. Meeresgesteine gleichen Alters lassen
sich zwar auf allen Kontinenten nachweisen, doch enthalten sie
nirgendwo außer in den unterdevonischen Hunsrückschiefern
die ersten, noch primitiven Ammoniten, deren Verbreitung sich vom
Hunsrück aus zudem verfolgen läßt auch bis in die
außereuropäischen Meere des Erdaltertums. Die
Paläontologie geht davon aus, daß die Ammoniten nach dem
Schlüpfen aus dem Ei zunächst ein gehäuseloses
Larvenstadium erlebten. Als frei im Wasser lebende Larven konnten sie
durch Meeresströmungen leicht verdriftet werden und eine
weltweite Verbreitung finden.

Medianschnitt
durch ein Cephalopodengehäuse (CYRTHOCERAS) aus dem
Mitteldevon der Eifel (FO: Sötenich). Erkennbar sind die
Wohnkammer, die daran anschließenden Luftkammern, die durch die
Kammerscheidewände (Septen) voneinander getrennt sind, und der
perlschnurartige Sipho, der alle Kammern miteinander verbindet.
(Länge des gekrümmten Gehäuses 20,5 cm.)
Foto:
Kreisbildstelle *)
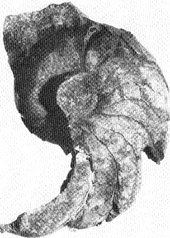
Gehäusefragment
eines zu den NAUTILOIDEN zählenden Kopffüßers
aus dem Oberen Mitteldevon des Bergischen Landes Bei Bergisch
Gladbach mit einem Durchmesser auf Nabelhöhe von 6,5 cm.
Foto:
Kreisbildstelle *)
Als Vorfahren der Ammoniten kommen
wohl zwei Tierformen infrage:
die Perlboote, die als
Vorläufer des heute noch vorkommenden NAUTILUS gelten,
die ausgestorbenen
Stabtintenfische (Bactriten).
Die Funde aus den unterdevonischen
Hunsrückschiefern lassen den Schluß zu, daß der
Übergang von den Bactriten zu den ersten Ammonitenformen „nur“
etwa fünf Millionen Jahre beansprucht hat. Obwohl es sich
hierbei um eine ungewöhnlich kurze Evolutionsphase handelt,
sprechen die fossilen Belegstücke jedoch gegen eine
Makromutation und für normale bilologische Übergänge
von einer Tiergruppe zur anderen.
Bei den Ammoniten ist die
Mannigfaltigkeit der Gehäuseformen und –verzierungen
besonders auffällig. Sie liefern wichtige Hinweise für die
Bestimmungen der Gattungen und Arten. Zu den bedeutsamsten
diagnostischen Merkmalen zählen vor allem die Lage des Siphos,
die Ausbildung der Lobenlinie, der Querschnitt der Windungen und die
Art ihrer Aufrollung.
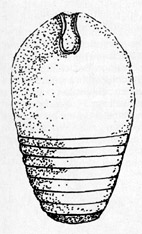
Ghomphoceras
Bohemicum
Darstellung eine vollständigen Gehäuses
von G. Bohemicum (BARR) aus dem Gotlandium der CSSR. G. Zählt zu
den Nautiloideen. Das birnenförmige, im Querschnitt kreisrunde
Gehäuse hat eine T-förmige Mündung und eng
aufeinanderfolgende Kammerscheidewände. Der zuweilen
perlschnurartig geformte Sipho liegt i. d. R. zentral. (Ca. 1½
nat. Größe.)
Wenn bei fossilen Muscheln und
Schnecken häufig nur Steinkerne als „Ausguß“ des
Schalen- oder Gehäuseinneren erhalten geblieben sind (als
„Füllung“ des Hohlraumes diente hierbei in der Regel
das umgebende Sediment), hat bei den Cephalopoden das Sediment
gewöhnlich nur Zutritt zur Wohnkammer. Die übrigen Kammern
sind durch die Scheidenwände hermetische abgeschlossen und darum
auch bei fossilen Exemplaren zumeist hohl oder / und mit Kristallen
ausgefüllt.
Andererseits kann aber durch den
Sipho oder durch beschädigte Partien der Schale Sediment auch in
die Luftkammern eindringen und „versteinern“.
Von der Devon- bis zur Kreidezeit
stellen Ammonoideen wichtige Leitfossilien. Für die Schnecken
und Muscheln gilt dies im Erdmittelalter (Mesozoikum) und in der
Erdneuzeit (Känozoikum).
An Beispielen mitteldevonischer
Cephalopoden sind neben anderen folgende Arten besonders
bemerkenswert:
Schnecken
Die Schnecken (Gastorpoda) sind
ähnlich formenreich wie die Kopffüßer, besitzen wie
diese auch einen im allgemeinen deutlich abgesetzten Kopf, der mit
Augen und paarig angeordneten stielförmigen Tentakeln als
Tastorganen (Fühlern) ausgestattet ist und an seiner Unterseite
die Mundöffnung zeigt. Im Mund befindet sich die Radula, eine
biegsame Raspelzunge, die mit reihenartig angeordneten spitzen
Zähnchen aus Chitin besetzt ist. Der oft sohlenartige, muskulöse
Fuß dient als Fortbewegungsorgan. Die Fußdrüse am
Vorderende des Fußes scheidet dazu einen Schleim aus, der die
Kriechfläche glättet. Der Sackförmige Weichtierkörper
enthält die Geschlechtsorgane, Herz, Leber, Niere und den Magen-
und Darmtrakt, der in den After mündet.
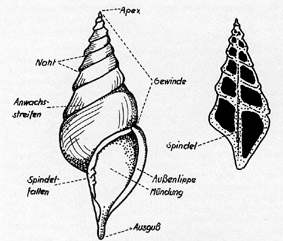
Rechtsgewundenes,
turmförmiges (trochispirales) Schneckengehäuse in Aufsicht
mit angedeuteter Skulptur; rechts im Längsschnitt (nach U.
Lehmann 1964).
Die Weichteile sind bei vielen
Schneckenarten durch ein Gehäuse geschützt. Ein
Spindelmuskel stellt die Verbindung zwischen dem Weichkörper und
dem einteiligen Schneckenhaus her, in das auch Kopf und Fuß
zurückgezogen werden können.
Wie bei den übrigen
Mollusken, wird auch das Gehäuse der Schnecken vom Mantel
ausgeschieden, der reich an Drüsen ist, die nicht nur den Kalk
für die Schale, sondern auch die Farbstoffe ausscheiden. Das
Schneckenhaus besteht in der Regel aus mehreren Windungen, die, wie
bei den planspiralen Formen, alle in einer Ebene liegen oder aber
trochispiral (turm- oder auch spindelförmig) mit mehr oder
weniger spitzem Wachstumswinkel aufsteigen.
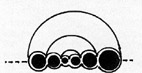
Beispiel
eines planspiralen Schneckengehäuses. Die Mittellinie aller
Windungen liegt in einer Ebene. Pseudoplanspirale Formen weichen nur
wenig von diesem Bauplan ab.
Die meisten Schneckengehäuse
sind rechtsgewunden, d. h. die Mündung liegt rechts. Es gibt
allerdings auch Schnecken, deren Gehäuse nicht eingerollt sind
(z. B. Napfschnecken). Die sogenannten Zahnschnecken (Dentalium)
besitzen ein röhrenförmiges, mäßig gebogenes und
an beiden Enden offenes Gehäuse.
Von besonderer Bedeutung für
die Bestimmung von rezenten oder auch fossilen Schnecken ist die je
spezifische Beschaffenheit der Außenseite des
Schneckengehäuses. Hier finden sich linienartige Vertiefungen,
Furchen oder Leisten, Rippen, Falten, auch Knoten oder sogar
Stacheln, die die Skulptur bestimmen. Sie werden als
Spiralverzierungen bezeichnet, wenn sie mit den Windungen verlaufen,
sie heißen Querverzierungen, wenn sie senkrecht oder schräg
zur Längsrichtung der Röhre angeordnet sind.
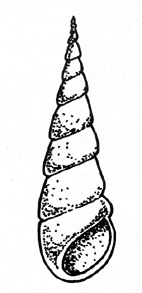
Ein
Trochispiral gewundenes Gehäuse. Die Windungen legen sich von
der Spitze an spiralig aneinander und werden von Umgang zu Umgang bis
zur Mündung stetig breiter. Ihr Umriß erreicht auf diese
Weise die Form eines Kegels.
Da das Schneckengehäuse,
ähnlich wie bei den Muscheln, eine enge Beziehung zu den
Weichteilen hat, ergeben sich selbst bei fossilen Schnecken
zusätzliche Bestimmungsmerkmale. So hat nämlich die
Organisation der Weichteile jeweils Spuren hinterlassen, die sich an
spezifischen Besonderheiten des Schneckengehäuses ablesen
lassen. Damit werden umgekehrt auch Schlüsse von versteinerten
Gehäusen auf die Struktur der Weichkörper fossiler
Schnecken hier und da je nach Erhaltungszustand der Funde durchaus
möglich.
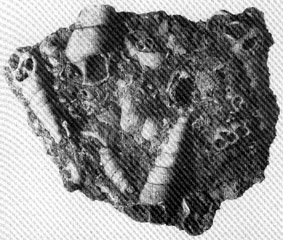
Eine
„Schneckenplatte“ mit TURITELLA aus dem Tertiär
von Ermingen bei Ulm.
(Länge der Platte 8,7 cm.)
Foto:
Kreisbildstelle *)
Schnecken existieren seit dem
frühen Erdaltertum. Sie treten bereits im Kambrium auf, bleiben
allerdings zunächst stammesgeschichtlich , ähnlich wie die
Muscheln, von untergeordneter Bedeutung. Während der Karbonzeit
zeigen sich allerdings neben Landschnecken auch die ersten
Süßwasserschnecken. Die marinen Formen bleiben den
devonischen Meeresschnecken noch sehr ähnlich. Erst mit dem
Erdmittelalter werden die Schnecken insgesamt häufiger und sind
dann in bestimmten Kalkgesteinen der Jura- und Kreidezeit
beträchtlich angereichert. Die Zunahme an Mannigfaltigkeit und
Verzierung beginnt bereits in der Trias und setzt sich dann in der
Jurazeit fort; verzierte Gehäuse herrschen hier nun ganz vor. In
der Tertiärzeit überflügeln die Schnecken die Muscheln
an Häufigkeit und erleben im Miozän (Jungtertiär) das
absolute Maximum ihrer Entwicklung.
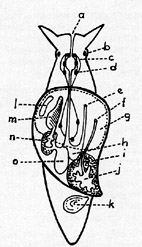
Schema
des Baues einer Schnecke (Ansicht von oben; das Gehäuse ist
weggenommen): a = Mund, b, c, d = Nervenknoten (Anhäufung von
Nervenzellen), e = After, f = Geschlechtsöffnung, g =
Mantelhöhle (Grenze durch punktierte Linie markiert), h = Magen,
i = Leber, j = Geschlechtsdrüse, k = Deckel, l = Geruchsorgan, m
= Kieme, n = Herz, o = Niere (Zeichnung nach Boas).
Auch heute sind Schnecken
außerordentlich zahlreich. Insgesamt sind ca. 85.000 Arten
bekannt. Ursprünglich existieren nur Meeresschnecken. In ihrer
langen Entwicklung haben sie aber sowohl das Süßwasser wie
auch das Festland als Lebensraum erobert. Die zum Landleben
übergegangenen Gastropoden mußten dabei die Kiemenatmung
zugunsten der Lungenatmung aufgeben.
Während die Süßwasser-
und Landschnecken ausnahmslos und von den Meeresschnecken viele Arten
Pflanzenfresser sind, leben andere Meeresschnecken ausschließlich
vom Plankton, wieder andere allerdings auch räuberisch. Diese
sind mit Hilfe ihrer scharfen Zähnchen in der Lage, zum Beispiel
Muschelschalen anzubohren und dann mit ihrem rüsselartigen Mund
in das Schaleninnere vorzudringen und ihr Opfer auszusaugen.

Straparollus
circinalis
S. weist zumeist ein trochispirale, hin und wieder
auch ein scheibenförmiges Gehäuse mit gerundeten, auf der
Oberseite leicht abgeflachten Umgängen auf. (Durchmesser auf
Nabelhöhe 8,0 cm.)
FO: Sötenich, Eifel,
Mittel-Devon.
Foto: Kreisbildstelle *)
Die Abbildungen zeigen einige
wenige Arten mitteldevonischer Schnecken.

Natica
sp.
Rundliches und zumeist glattes Gehäuse mit relativ
kleinem Gewinde. Die Endwindung ist i. d. R. groß. N. zählt
zu den „Bohrschnecken“. Die Bohrung erfolgt chemisch durch
Ätzung über eine sog. Bohrdrüse. (Durchmesser des
größten Gehäuses 2,0 cm.)
FO: Gondelsheim, Eifel,
Mittel-Devon.
Die Bestimmung devonischer
Schnecken ist oft schwierig, weil sehr häufig nur die Steinkerne
gefunden werden. Die Schalen sind oft nicht mehr erhalten. Damit
fehlen auch Hinweise auf die einstigen Strukturen, wie
Anwachsstreifen, Ziernähte und dgl.
Leider steht eine neue
wissenschaftliche Bearbeitung der mitteldevonischen Schnecken noch
aus.

Murchisonia
Archiace
Murchisonia kommt im Mittel-Devon des Rheinischen
Schiefergebirges in mehreren Arten vor (M. bilineata, M. tricineta,
M. pagodeformis, M. turbinata, M. coronata, M. bicoronata, u. a.,
nach W. Paeckelmann, 1922).
Das nicht sehr große, schlanke,
turmförmige (trochispirale) Gehäuse besteht aus zahlreichen
Umgängen. Die äußere Skulptur weist je nach Art
Längskiele oder –streifen, Rippen oder Knötchen auf,
bei einigen Arten in doppelter Anordnung. Die ausgußartige
Mündung hat die Form einer Rinne, ihr Außenrand ist
scharf. Die Abb. zeigt links Murchisonia Archiaci var. turbinata
(4,5 cm lang), FO: Bergisch Gladbach, und rechts Murchisonia
Archiaci var. bilineata. FO: Bergisch Gladbach, Schlade, Bücheler
Schichten, Ob. Mittel-Devon. M. bilineata zeigt eine gerundete,
wulstige Kante unter der Naht. Die Windungen haben ein schwach
konkaves Profil. Die Art ist stellenweise häufig
anzutreffen.
Foto: Kreisbildstelle *)

Euromphalus
laevis
Das planspirale, scheibenförmige Gehäuse hat
einen weit geöffneten Naben und auf der Oberseite einen schmalen
Kielwulst. Alle Umgänge berühren sich. Die Mündung des
Gehäuses ist kreisrund. Das abgebildete Exemplar hat auf
Nabelhöhe einen Durchmesser von 2,0 cm.
Euomphalus reicht vom
Gotlandium bis ins Mittl. Perm und tritt vor allem im Karbon weltweit
mit zahlreichen Arten auf.

Macrochilina
arculata
Eine große Schnecke mit treppenförmig
abgesetzten Windungen, die sich nach oben relativ rasch verjüngen.
Die Windungen zeigen einen stark entwickelten Wulst und kräftige
Anwachsstreifen. Die Höhe des abgebildeten kegelförmigen
Gehäuses beträgt 8,9 cm.
FO: Kombüchen,
Herrenstrunden, Bergisches Land, Mittel-Devon.
Foto:
Kreisbildstelle *)

Loxonema
sp.
Das Gehäuse ist schlank, hoch getürmt. Es hat
zahlreiche und nur wenig gewölbte, glatte Umgänge. (Höhe
3.3 cm.)
FO: Gondelsheim, Eifel, Mittel-Devon.

Pleurotomaria
delphinuloides
Das Gehäuse ist mittelgroß, kegel-
oder kreiselförmig, seine Oberfläche spiralig gestreift.
Die Windungen sind gebläht. Der Nabel ist tief, trichterförmig.
(Breite des Gehäuses 2,8 cm.)
FO: Bergisch Gladbach, Schlade,
Massenkalk, Bücheler Schichten.

Bellerephon
striatus
Diese nicht allzu häufig mehr vorkommende
Schneckenart kann eine ansehnliche Größe erreichen. Der
Durchmesser des vorliegenden, besonders schönen Exemplars
beträgt auf Nabelhöhe 6,3 cm. Der Nabel selbst ist
normalerweise eng und auch tief. Über das Gehäuse mit den
kräftigen Rippen läuft ein deutlich hervorstehender Kiel.
Die Umgänge sind breit gerundet.
FO: Bergisch
Gladbach-Büchel.
Foto: Kreisbildstelle *)
Tentaculiten
Die Tentaculiten werden, obwohl
ihre Stellung im zoologischen System noch umstritten ist, neuerdings
als eigene Klasse innerhalb des Stammes der Weichtiere aufgefaßt.
Es handelt sich um marine Organismen mit einem kleinen,
spitzkegelförmigen, kalkigen Gehäuse, dessen Anfangsteil
gekammert ist. Seine Oberfläche zeigt parallel-querverlaufende,
engständige Ringe. Wahrscheinlich besaßen die Tentaculiten
Fangarme.

Eine
Gesteinsplatte (9,5 cm lang) mit „eingeregelten“
TENTACULITEN.
Rechts: Längsschnitt durch ein
Tentaculitengehäuse, teilweise mit Wiedergabe der Skulptur
Tentaculiten erscheinen im
Ordovicium und treten während der Devonzeit stellenweise in
bemerkenswerter Häufigkeit auf, wobei sie im Mitteldevon ganze
Schichtflächen bedecken können. Im Oberdevon sterben sie
bereits wieder aus.
Fotos: Verfasser und
Kreisbildstelle, Reproduktionen Heinz-Josef Weingarten und Jürgen
Reiher *)
Entnommen: Kreis Euskirchen -
Jahrbuch 1979/80
© Copyright
wisoveg.de 2003
©
Copyright 2003
Kreisarchiv
Zu
den Euskirchener Wisoveg-Seiten
Zur
Homepage